Від тіла нейрона відходить багато відростків, або нейритів. Аксони - це відростки, що несуть електричні імпульси від тіла клітини, тоді як дендрити сприймають імпульси (проводять їх до тіла нейрона). Аксони проводять імпульси лише в одному напрямку. Деякі з них сягають довжини більш як 1м, причому аксони з великим діаметром проводять імпульси дуже швидко.
Нервова клітина з усіма її відростками вкрита дуже тонкою білково-ліпоїдною оболонкою.
Відростки нервових клітин називаються нервовими волокнами (рис. 24). Виходячи за межі мозку, вони утворюють своїм скупченням нерви. Нерв завжди вкритий сполучнотканинною оболонкою.
Нервові волокна вкриті клітинами, які було відкрито ще Шванном і які так і називаються шваннівськими клітинами; всі разом вони утворюють шваннівську оболонку. Ці клітини розташовані, як суцільний ланцюг, уздовж волокна. Вони мають значення в обміні нервового волокна і виконують роль ізолятора. Навколо одних волокон під шваннівською оболонкою утворюється досить товстий шар мієліну - речовини ліпоїдної природи, навколо інших мієлінової оболонки немає. Залежно від цього волокна поділяються на мієлінові і ті, які не мають мієліну.
Більшість нервів, які іннервують тіло (м'язи, зв'язки, сухожилки, надкісницю кісток), є мієліновими. Мієлін має жовтуватий колір, а мієлінова оболонка через проміжки рівної довжини переривається, залишаючи відкритими ділянки осьового циліндра. Це вузли нервового волокна (перехвати Ранв'є).
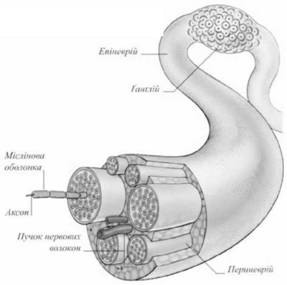
Рис. 24. Будова нервового волокна (за Дорлінг Кіндерслі, 2003)
Немієлінові волокна не мають мієлінової оболонки, ізольовані один від одного тоненькою ендотеліальною оболонкою і зустрічаються переважно в нервах автономної нервової системи.
Безмієлінові та мієлінові нервові волокна відрізняються своєю товщиною і особливістю проведення нервових імпульсів. Безмієлінові волокна мають діаметр 1-4 мкм, входять до складу вегетативних нервів і проводять нервові імпульси зі швидкістю 1-2 м/с. Мієлінові волокна товстіші, діаметр їх коливається від 1 до 20 мкм, швидкість передачі нервових імпульсів значно вища - 5-120 м/с. Вони зустрічаються переважно у складі периферійних нервів, а в центральній нервовій системі мієлінові волокна мають дещо іншу будову: їхню оболонку утворюють олігодендроцити, в яких відсутні насічки мієліну, а вузлові перетяжки більші за розмірами.
Периферійні нерви, або нервові стовбури складаються з нервових волокон (мієлінових і безмієлінових) і сполучнотканинних оболонок. Між нервовими волокнами в складі нерва знаходяться тонкі сполучнотканинні прошарки - ендоневрій. Група нервових волокон утворює нервовий пучок, оточений товстішою оболонкою зі сполучної тканини, яка називається периферією. Нервові пучки об'єднуються в нервовий стовбур, або периферійний нерв, вкритий зверху епіневрієм - оболонкою зі сполучної тканини, багатої фібробластами, макрофагами і жировими клітинами. Сполучнотканинні оболонки нерва містять кровоносні і лімфатичні судини та нервові закінчення.
6. Властивості нервової системи
Основними властивостями нервової системи є збудливість, провідність і гальмування.
Збудливість проявляється у виникненні збудження у відповідь на дію того чи іншого подразника. Здатність живих систем під впливом подразників переходити із стану фізіологічного спокою до стану активності називається подразливістю.
Подразники організму поділяють на внутрішні і зовнішні. Внутрішні подразники - це фізичні і хімічні зміни внутрішнього середовища, наприклад, зміна осмотичного тиску, хімічного складу крові, вплив гормону, вуглекислого газу, нервового імпульсу.
Зовнішні (контактні і дистантні) подразники бувають трьох видів:
o фізичні - механічні (тиск, укол), температурні, звукові, світлові, електричні;
o хімічні - кислоти, луги, солі, пахучі і смакові речовини, отрути, тощо;
o біологічні - всі живі істоти, віруси.
За фізіологічним значенням розрізняють адекватні і неадекватні подразники. Адекватний подразник - агент, що діє на тканину, пристосовану для його сприймання (наприклад, світло - на око, нервовий імпульс на м'яз). Усі інші подразники для цієї тканини неадекватні.
Найменша сила подразнення, здатна викликати збудження, називається пороговою силою. Сила подразнення, яка не викликає збудження, називається підпороговою. Сила подразнення, більша за порогову, називається надпороговою.
Виникнення і поширення збудження пов'язані зі зміною електричного заряду живої тканини, з так званими біоелектричними явищами. Між зовнішньою поверхнею клітини і її цитоплазмою у стані спокою створюється різниця потенціалів (близько 60-90 мВ), причому поверхня клітини заряджена електропозитивно щодо цитоплазми. Ця різниця потенціалів називається потенціалом спокою. Величина для клітин нервової і м'язової тканини становить 80-90 мВ, для епітеліальної 18-20 мВ. Причиною такої різниці потенціалів є вибіркова проникність мембрани.
Мембрана складається з ліпідів, білків і глікозаміногліканів. Біомолекулярний шар ліпідів є матриксом, в який занурені білкові молекули, що утворюють канали для води й іонів, формують йонні насоси. Глікозаміноглікани розташовані на поверхні мембрани і виконують рецептивні Функції (рис. 25).
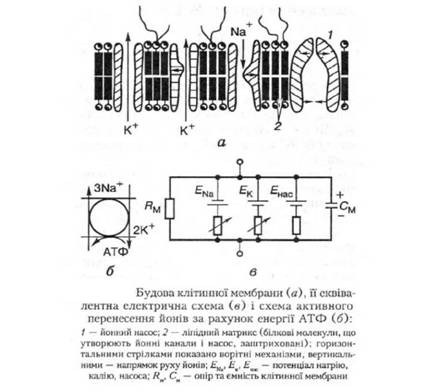
Рис. 25. (за Чайченко П. М., Цибенко В. О., Сокун В. Д., 2003)
Клітинна мембрана має значний опір, який залежить від стану її йонних каналів і ємності, яка створюється її ліпідним матриксом, і є досить сталою.
Клітинна мембрана легко проникна для жиророзчинних речовин, молекули яких проходять через її ліпідний матрикс. Водорозчинні молекули великого діаметра, у тому числі й аніони органічних кислот, практично не проникають крізь мембрану.
У стані фізіологічного спокою клітинна мембрана є проникною для К , оскільки в ній є спеціальні калієві канали, що пропускають лише гідратований калій.
Усередині клітини в цитоплазмі в 30-50 разів більше іонів калію, у 8-10 разів менше іонів натрію і в 50 разів менше іонів хлору, ніж на поверхні клітини. У стані спокою клітинна мембрана більш проникна для іонів калію, ніж для іонів натрію. У стані спокою нейрона, коли імпульс не передається, позитивно заряджені іони калію дифундують зсередини клітини з постійною швидкістю. Таким чином, внутрішня поверхня стає негативно зарядженою за рахунок іонів хлору, амінокислот та інших іонів.
Мембранний потенціал спокою (МПС) створює електричне поле значної напруги - 10 В/см . Це поле діє на макромолекули мембрани й надає їх зарядженим групам певної просторової орієнтації. Електричне поле МПС забезпечує функцію йонних каналів клітинної мембрани.
Той факт, що в умовах спокою № постійно надходить у клітину, а К виходить з неї має дуже важливі наслідки, оскільки система не може бути фактично врівноважена за допомогою звичайної дифузії і простого відновлення заряду на клітинній мембрані. Якщо немає інших процесів, то клітина дуже швидко накопичуватиме № і втрачатиме К , що зменшить їх трансмембранний градієнт. Проте такого не відбудеться, скільки цьому протидіє натрій-калієвий насос. Він працює споживаючи енергію АТФ, і його основним компонентом є фермент мембранна натрій-калій-АТФ-аза.
Для збудження нейрона подразник повинен перетворити заряд усередині клітини з негативного на позитивний. Рівень, на якому стимул починає передавати електричний імпульс називається порогом. Якщо подразник слабкий і не сягає порога, у мембрані виникає короткочасна місцева реакція. Однак, якщо поріг досягнутий, імпульс проходить по всій довжині волокна. Швидкість проведення може бути різною: охолоджені волокна (якщо прикладають лід для послаблення болю), волокна малого діаметра та волокна без мієлінових оболонок проводять імпульси повільніше.
У разі стимуляції нервовим імпульсом позитивно заряджені іони з позаклітинної рідини проникають у мембрану клітини. У цих місцях електричний заряд на внутрішній поверхні клітинної мембрани змінюється з негативного на позитивний. Ця локалізована зміна заряду через мембрану стимулює подібні зміни у наступних сегментах мембрани. Електричний імпульс продовжує рухатись вниз по аксону; при цьому попередні сегменти мембрани повертаються до положення "внутрішньо-негативного" стану. Імпульс досягає синаптичної щілини. Медіатор, вивільнившись, проходить через синаптичну щілину і стимулює скорочення м'язових волокон.
Якщо на ділянку нервового або м'язового волокна діяти сильним подразником, то в ній виникає збудження, яке виявляється в швидкому коливанні мембранного потенціалу і яке називається потенціалом дії. Причина виникнення потенціалу дії - зміна іонної проникності мембрани. При подразненні іони натрію надходять всередину клітини, тому що вони заряджені позитивно і їх тягнуть всередину електростатичної сили і тому, що їх концентрація всередині клітини невелика. При цьому потік позитивно заряджених іонів натрію із зовнішнього середовища перевищує потік іонів калію із клітини назовні. В результаті внутрішня поверхня мембрани стала заряджена позитивно, а зовнішня негативно. В цей момент реєструється потік потенціалу дії, коли деполяризація мембрани досягає критичного рівня. Підвищення проникності мембрани для іонів натрію триває недовго. Слідом починаються в клітині відповідні процеси, проникність мембрани для іонів натрію знижується, а для іонів калію зростає. Активний транспорт іонів калію представлений ферментною системою в мембрані нервового волокна. Активується ця система іонами натрію, калію, магнію, джерелом енергії є АТФ.
Натрієва провідність досягає максимуму менш ніж за 1мс (мілісекунд) після початку ПД, а потім різко знижується, зникаючи зовсім протягом 0,5мс, хоча деполяризація мембрани ще зберігається. Це явище називають натрієвою інактивацією, яка існує доти, доки триває деполяризація мембрани. Натрієва активаційна система відновлюється, коли МПС повертається до початкового рівня або навіть при певній гіперполяризації мембрани.
Під час гальмування чи блокування електричних імпульсів, канали, чутливі до іонів хлору або калію, можуть відкриватись швидше, ніж канали, чутливі до натрію. Позитивні іони калію виходять з клітини-мішені або ж негативні іони хлору проникають у мембрану клітини. В обох випадках електричний заряд на внутрішній поверхні мембрани клітини-мішені залишається негативним, нейрон не збуджується і нервовий імпульс гальмується.
Провідність - здатність передавати збудження, що виникло, - є другою важливою властивістю нервової тканини. Проведення збудження можливе лише за умови цілості нерва і збереження його життєвих властивостей. В умовах цілого організму всі імпульси в нервовій системі проводяться лише в одному напрямку. Пояснюється це тим, що контакти між сусідніми нейронами, так звані синапси (від грец. synapsis - зв'язок), проводять збудження лише в напрямі від доцентрового нейрона на відцентровий і нездатні проводити його в зворотному напрямі (рис. 26).
За структурою та локалізацією синапси поділяють на такі групи: міжнейронні, рецепторно-нейронні та нейро-ефекторні.
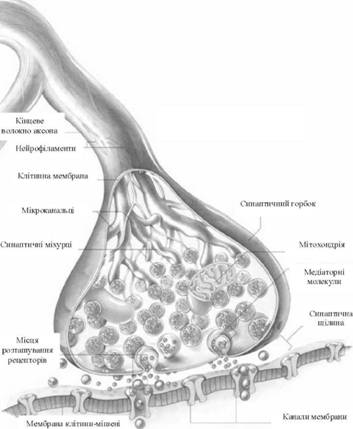
Рис. 26. Будова синапса (за Дорлінг Кіндерслі, 2003)
Термін синапс увів у науку Ч.Шеррінгтон (1897) для позначення функціонального зв'язку між нейронами. Залежно від будови і функціонування розрізняють хімічні та електричні синапси.
Міжклітинна передача імпульсів здійснюється за допомогою трьох типів міжклітинних контактів:
а) хімічних синапсів (збудження і гальмівні впливи передаються в цих синапсах за допомогою медіаторів);
б) електричних синапсів (вони функціонують за рахунок ионного струму);
в) нексусів, або щільних контактів (властивий низький опір і висока двобічна проникність для йонів).
Синапс утворений двома мембранами - пресинаптичною, яка знаходиться на нервовому закінченні і має вигляд ґудзиків, кілець, бляшок, і постсинаптичною, яка міститься на тілі або дендритах нейрона, до якого передається нервовий імпульс. Збудження через синапси передається хімічним шляхом за допомогою медіатора, який міститься в синаптичних міхурцях, що розташовані в синаптичній бляшці. Найчастіше виробляються серед медіаторів: ацетилхолін, адреналін і норадреналін, рідше серотонін, речовина Р, глутамінова кислота, енкефалін, нейротензин, ангіотензин II, дофамін, гліцин та ін.
Ацетилхолін - медіатор нервово-м'язових синапсів, ряду типів синапсів у вегетативній (автономній) нервовій системі і в ЦНС. Ацетилхолін накопичується у синаптичних пухирцях. Один нервовий імпульс спонукає вивільнення вмісту 100-300 пухирців. Токсин ботулізму (білок) гальмує пресинаптичне вивільнення ацетилхоліну в холінергічних синапсах.
Катехоламіни накопичуються у синаптичних пухирцях (10-15 тис. молекул на один пухирець). При надходженні нервового імпульсу катехоламіни вивільняються із пухирців шляхом екзоцитозу в синаптичну щілину і зв'язуються з рецепторами постсинаптичної мембрани.
Норадреналін виконує роль медіатора у нервових закінченнях постгангліонарних синаптичних волокон і у синапсах різних відділів ЦНС, причому може бути збуджувальним і гальмівним медіатором, що зв'язано з типом адренорецептора.
Дофамін є нейромедіатором базальних гангліїв головного мозку, зокрема в нейронах, які зв'язують чорну речовину зі смугастим тілом і беруть участь в регуляції рухів. Інші дофамінергічні системи мозку відіграють роль у здійсненні інтегративних функцій мозку, у процесах відчуття, емоцій, пам'яті.
Дофамінергічна передача імпульсів порушується при ураженні чорної речовини і смугастого тіла - хворобі Паркінсона, яка характеризується регідністю м'язів, тремором і гіпокінезією. У хворих знижена кількість дофаміну і продуктів його розпаду.
Серотонінергічні нейрони беруть участь у регуляції сну, настрою, емоцій, відчутті болю. Синтезується серотонін із триптофану.
Гальмівним медіатором ЦНС служить гамма-аміномасляна кислота (ГАМК). Синтезується вона із глутамату.
Відкриті ГАМКд- ГАМКв - рецептори. Перші частіше зустрічаються в мозочку, корі головного мозку і гіпоталамусі, а другі рівномірно розподіляються по всьому мозку.
Амінокислота гліцин також гальмівний медіатор ЦНС, зокрема в спинному мозку і в стовбурі головного мозку. Гліцин підвищує проникність постсинаптичної мембрани для СІ-, визначаючи її гіперполяриза-цію і гальмуючи діяльність нейронів.
Постсинаптичне гальмування гліцином припиняється при введенні алкалоїду стрихніну, який зв'язується, вірогідно, з іонним каналом.
В гіпокампі та інших відділах відкрито більше десяти типів рецепторів для глутамату. Кількість рецепторів збільшується при стимуляції.
Припускають, що глутамат бере участь у тривалих синаптичних змінах, пов'язаних з навчанням і пам'яттю.
У 1970-х рр. встановлено, що властивості нейромедіаторів проявляють низькомолекулярні пептиди-енкефаліни. Вони зв'язуються з опіатними рецепторами, тобто з рецепторами морфіну (алкалоїду з опійного маку) та його аналогів, і тому їх називають ендогенними опіатами. Як і морфін, опіатні нейропептиди проявляють знеболювальну і ейфоричну дію. Крім енкефалінів, до групи опіатних пептидів відносяться ендорфіни. Опіатні рецептори і нейропептиди знайдені в багатьох ділянках мозку, зокрема в тих, які відповідають за проведення сигналів болю і за регуляцію емоцій.
Надходження нервового імпульсу в пресинаптичне закінчення супроводжується викиданням в синаптичну щілину, яка знаходиться між мембранами, медіатору із синаптичних міхурців. Чим більша сила подразнення, тим більше виділяється медіатора в синаптичну щілину, розміри якої дуже малі, і медіатор швидко досягає постсинаптичної мембрани, взаємодіючи з її речовиною. В результаті цієї взаємодії проникність для іонів натрію підвищується, що веде до переміщення іонів, і, як наслідок, виникає збудливий постсинаптичний потенціал, виникає збудження, яке поширюється. Через кілька мілісекунд медіатор руйнується спеціальними ферментами. Вважають, що в спеціалізованих гальмуючих нейронах, у нервових закінченнях аксонів виробляється особливий медіатор, який діє гальмуюче на наступний нейрон. У корі головного мозку таким медіатором вважають у-аміномасляну кислоту. Медіатор гальмування, взаємодіючи з речовиною постсинаптичної мембрани, збільшує її проникність для іонів калію і хлору. В результаті відбувається не зниження величини внутрішнього заряду мембрани, а підвищення внутрішнього заряду постсинаптичної мембрани, відбувається її гіперполяризація, в результаті чого настає гальмування.
Проведення збудження зумовлене тим, що потенціал дії, який виник в одній клітині чи в одній з її ділянок, стає подразником, що спричиняє збудження сусідніх ділянок. Збудження в мієлінових волокнах виникає тільки в тих ділянках, які не вкриті мієліновою оболонкою, у вузлах нервового волокна і поширюється по них стрибкоподібно (120 м/сек). По немієлінових нервових волокнах збудження поширюється повільно (від 1 до 30 м/сек). Це пов'язано з тим, що іонні процеси, які відбуваються через мембрану волокна, проходять по всій довжині волокна. Збудження від однієї нервової клітини до іншої передається з аксона одного нейрона на тіло клітини і дендрити нейрона.
Крім процесів збудження, в діяльності ЦНС важливу роль відіграють і процеси гальмування, які є своєрідним діяльним станом, що викликаний збудженням і з ним пов'язаний. Гальмування запобігає виснаженню нервових клітин при дуже сильних і частих подразненнях. Розрізняють пресинаптичне, постсинаптичне і вторинне гальмування. Пресинаптичне гальмування розвивається в пресинаптичних розгалужених аферентних аксонів, завдяки чому блокується проведення імпульсів до синапсів і виникає гальмування реакції відповіді. При постсинаптичному гальмуванні імпульс, який прийшов до гальмівного синапсу, зумовлює гіперполяризацію постсинаптичної мембрани. При цьому зростає величина мембранного потенціалу і виникає гальмівний постсинаптичний потенціал, в результаті чого настає гальмування. Вторинне гальмування здійснюється без участі спеціальних гальмівних структур і розвивається в збудливих синапсах при дії подразників надмірної сили.
Численними дослідами встановлено, що збудження в одних центрах нервової системи викликає гальмування в інших. Наприклад, коли м'язи-згиначі правої ноги і розгиначі лівої скорочуються, розгиначі правої ноги і згиначі лівої перебувають у розслабленому стані. Така координована діяльність м'язів нижніх кінцівок пояснюється тим, що збудження в нервових центрах згиначів викликає гальмівний стан в центрах цієї ж кінцівки. При цьому збудження в центрі згиначів однієї кінцівки зв'язане з гальмуванням у центрі згиначів другої. Згідно з ученням видатного російського фізіолога М.Е. Введенського, гальмування і збудження є лише два прояви одного і того самого нервового процесу, які залежать лише від сили і частоти подразнення.
Периферичні нервові волокна, розплющені або частково перерізані, повільно регенерують, якщо тіло клітини і сегменти мієлінової оболонки неперервні. Регенерація волокон не відбувається в головному та спинному мозку. Ушкоджені нервові волокна трансформуються в тканину рубця та іннактивуються.
Якщо нервове волокно ушкоджується в місці, найбільш віддаленому від тіла клітини, настає дегенерація внаслідок нестачі життєво важливих білків та ензимів, і мієлінова оболонка стає порожнистою. Неушкоджене тіло нейрона стимулює ріст декількох нервових відростків у залишках волокна. Один з цих відростків може прорости через порожню, але неушкоджену мієлінову оболонку. Нове нервове волокно, яке росте зі швидкістю близько 1,5 мм на день, досягає попереднього місця розташування, функція і відчуття повільно відновлюються, а невикористані нервові відростки дегенерують.
Дія стрихніну та наркозу на збудливість нервової системи
Центральна нервова система має специфічну чутливість до деяких речовин, зокрема до стрихніну та хлороформу. Стрихнін підвищує збудливість синапсів нервових клітин спинного мозку настільки, що після введення цього препарату навіть у незначних дозах підпорогові подразнення викликають збудження, яке поширюється (іррадіює) і дає судорожні скорочення м'язів тулуба. Жаба, отруєна стрихніном, спершу пересувається, широко розкинувши кінцівки, потім тонус м'язів підвищується, лапки жаби стають ніби дерев'яними, слабке доторкування до підставки, на якій лежить жаба, викликає загальні тетанічні судороги. Подразнення у такої тварини викликає не координовану рефлекторну реакцію, а загальні судороги.
Протилежний ефект має дія хлороформу. Ця речовина викликає загальне порушення обміну речовин у клітині, але найбільш чутливі до неї нервові клітини мозку. Хлороформ викликає загальне зниження чутливості, аж до повного наркозу. Жаба, отруєна хлороформом, не реагує на подразнення, тонус м'язів у неї майже зовсім зникає.
8. Поняття про рефлекс. Рефлекторна дуга
9. Основні етапи розвитку нервової системи
З історії науки
Еволюційний процес
ЛЕКЦІЯ 3. Будова, функції та розвиток спинного мозку
1. Будова спинного мозку
1.1. Оболонки спинного мозку
1.2. Лікворна система мозку
1.3. Поперечний переріз спинного мозку