У трофічних ланцюгах всі види речовин послідовно переходять від одного організму до іншого. Органічні речовини в цьому процесі перетворюються в специфічну форму для кожного виду рослин та тварин. Так, білки рослин в процесі живлення фітофагів, які їх споживають, розщеплюються до амінокислот, і вже з них в організмі тварини синтезують свої специфічні білки.
Інша доля притаманна окремим хімічним речовинам, зокрема так званим ксенобіотикам - речовинам, що в природі спочатку були відсутні, а потім синтезовані людиною. Такі речовини проходять через трофічні ланцюги незмінними. У силу того, що розмір біомаси в екологічних пірамідах закономірно знижується при переході на кожний новий трофічний рівень, концентрація ксенобіотиків у розрахунку на одиницю біомаси стає більшою. Цей ефект називається законом концентрування речовин у трофічних ланцюгах.
Концентрування речовин у трофічних ланцюгах має важливі наслідки для всієї практики господарювання людини в природних екосистемах. Забруднення, яке вважається незначним при оцінці кількості забруднювача в навколишньому середовищі, стає катастрофічно небезпечним при дії закону концентрування та небезпечним, перш за все, для самої людини, яка знаходиться на вершині усіх трофічних ланцюгів пасовищного типу.
Трофічні ланцюги виконують ще й бар'єрну функцію, із концентруючими та бар'єрними функціями живої речовини пов'язана здатність екосистем до самоочищення вона проявляється стосовно великого класу речовин. Ряд з них, потрапляючи до трофічного ланцюга, поступово руйнується. Але така здатність біномів до самоочищення не безмежна. Є верхній граничний рівень концентрації, перевищення якого вже не дає можливості біому очиститися від даної речовини. Тут багато чого залежить від типу забруднюючої речовини та швидкості її надходження в екосистему. При поступовому надходженні забруднюючих речовин самоочищення йде ефективніше, ніж при разових викидах в екосистему великої кількості ксенобіотиків.
Деякі забруднюючі речовини в біогеохімічних циклах не руйнуються, а переходять у депо циклу (гірські породи, атмосферу і т.п.), та їхня шкідлива дія на організм знижується. Це відбувається з багатьма мінеральними речовинами.
2.8. Розвиток та еволюція екосистем
Екосистемам, як і усім природним об'єктам, властиві закономірні зміни у часі. Ці зміни відповідають принципу самостійного розвитку та руху матерії. Для позначення явища розвитку екосистем звичайно використовують термін "еволюція". Він не зовсім вдалий. Зі словом "еволюція" у більшості випадків асоціюється історичний розвиток, боротьба за існування та природний добір. На рівні екосистем цих явищ немає, але термін "еволюція" тут застосовується більшістю авторів і змінювати його немає підстав. Хоча, за М.А. Голубцем (1969), доводиться говорити про екосистемну еволюцію. Екосистемна еволюція включає в себе "зміну у часі просторової та функціональної організації екосистем" (М. Голубець, 1982).
В екосистемній еволюції існує дві узгодженні форми розвитку:
a) еволюція живих організмів;
b) самоорганізація неживої матерії.
Закономірності біологічної еволюції, відкриті Ч. Дарвіним, вже добре вивчені. Порівняно новою сторінкою в екології є встановлення фактів та механізмів самоорганізації в неорганічному світі.
Відкриття самоорганізації в неорганічному світі як противаги зростанню ентропії має епохальне значення, і його філософські наслідки ще до кінця неусвідомлень
У неживій матерії до саморозвитку здатні відкриті системи, що складаються з підсистем із колективною поведінкою. На противагу до другого закону термодинаміки, в неорганічній матерії, однак, відомі випадки, коли порядок виникає з безладу.
В екології розвиток у формі самоорганізації матерії є важливою властивістю екосистем. Кооперативна поведінка живої матерії, яка еволюціонує, та абіотичних компонентів екосистем, що самоорганізуються, веде до виникнення все нових і нових форм організації. Вони і складають зміст екосистемної еволюції. Відкритий характер екосистем зумовлює те, що їхня еволюція визначається внутрішніми особливостями екосистем, які само-розвиваються. Але вона здійснюється також і за рахунок зовнішніх стосовно екосистем збурень.
Внутрішнім джерелом самоорганізації екосистем є протиріччя форм та темпів розвитку живого та неживого компонента, їхньої структури. Детальний аналіз, проведений М.А. Голубцем (1982), показав, що за рахунок еволюції екосистем у біосферний кругообіг речовин включаються все нові потенційні середовища життя і підвищується продуктивність та стабільність біогеоценотичного покриву Землі в цілому. Ще до нього А.І. Літока (1925) сформулював правило максимуму потоку енергії в біологічних системах, відповідно до якого екосистемна еволюція зорієнтована так, що все більша й більша частка енергії направляється на збільшення незалежності та автономності екосистем щодо зовнішніх збурень.
Важливим рушієм поступових змін екосистем є процес життєдіяльності організмів. За повної замкненості біогеохімічних циклів в екосистемах накопичуються органічні та неорганічні залишки специфічного характеру. Так, очевидно, що на самих ранніх етапах еволюції екосистем в них не було організмів, які використовували для дихання вільний кисень тому, що його в атмосфері просто не було. У міру накопичення кисню, в результаті фотосинтезу зелених рослин, на Землі почали формуватися екосистеми, що вміщували живі організми, які дихали киснем.
Інший рушій екосистемної еволюції - це сама біологічна еволюція, тобто зміна організмів різного рівня організації. Як показано М.А. Голубцем, вся сукупність генотипів тієї чи іншої екосистеми, що складає генопласт, також є об'єктом еволюції. Ю Одум (1986) підкреслював важливість для еволюції екосистем двох ефектів: коеволюції та групового добору. Поява тісної кооперації типу "рослина та її фітофаги", "жертва та хижак , спеціалізовані квітки та їх запилювачі - це все результат коеволюції.
Дані геології свідчать, що еволюція екосистем зумовлена ходом вікових змін самої геоморфологічної структури Земної Кулі та пов'язаними з ними змінами клімату. Аналізуючи фактори еволюції екосистем, М.А. Голубець (1983) підкреслював, що екосистеми як природні структури утримують значну кількість акумульованої в них вільної енергії. Це спричиняє їхній нестійкий стан та веде до періодичних самозбурень. Таким чином, можна стверджувати, що процес екосистемної еволюції базується на трьох основних факторах:
1)зміні середовища;
2)спадковій мінливості живих організмів та природному доборі;
3)наявності в екосистемах вільної енергії.
Загальний аналіз закономірностей зміни екосистем показує, що в умовах більш чи менш стабільного екологічного середовища екосистеми, які мають велике внутрішнє різноманіття, витісняють прості екосистеми. Екосистемна еволюція йде від простого до складного. За B.C. Голубєвим (1992), критерієм прогресивності еволюції екосистем та біосфери є темп нарощування в них вільної енергії, що сприяє збільшенню стійкості функціонування екосистем та здатності до саморегуляції.
Динаміку розвитку великих екосистем та біосфери в цілому можна побачити з аналізу окремого випадку еволюції екосистем -сукцесії.
Сукцесія - це послідовна зміна біогеоценозів (екосистем) на одній і тій же території під впливом природних факторів або діяльності людини.
Початковою точкою такого розвитку при розгляді сукцесії зручно вважати такі ділянки, як повністю не зайняті будь-якими живими організмами. Це, наприклад, поля лави, оголення ґрунту після зсуву, свіжі річкові наноси. Угрупування живих організмів, які першими опиняються в таких місцях, називаються піонерами. У міру розвитку будь-якого піонерного угрупування, воно рано чи пізно досягає стійкої рівноваги, коли угрупування не може замінюватися іншим угрупуванням. Такі угрупування називаються клімаксними.
Повний набір угрупувань живих організмів у часовій послідовності їхніх змін від піонерного до клімаксного складає сукцесій ний ряд.
Аналізуючи екосистеми, розрізняють сукцесію екосистеми в цілому та окремо сукцесії рослинності, тваринного та мікробного населення. У типовому випадку узагальнена схема сукцесії полягає в послідовній зміні нижчих рослин і тварин більш організованими, а в рослин - ще й багаторічними формами (рис.2.6.). Розрізняють первинні сукцесії, коли заселяється початково позбавлений життя субстрат, та вторинні сукцесії, коли формування екосистеми йде на базі такого угрупування, що раніше існувало, але згодом було зруйнованим. Вторинні сукцесії беруть початок на місцях згарищ, вирубок, занепалого сільськогосподарського землекористування і т.п. Концепція сукцесій була сформована в 1916 році Ф.Е. Клементсом. На його думку, всі основи і сукцесії обов'язково завершуються одним клімаксним угрупуванням, яке відповідає даному клімату. Такий підхід отримав назву моноклімаксу.
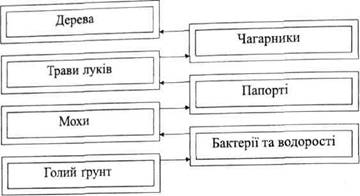 |
Рис. 2.6. Принципова схема сукцесії в лісовій зоні
Пізніше було показано, що й в умовах одного типу клімату, залежно від характеру ґрунту, гідрологічного режиму і т.п., формується цілий набір різних, але стійких угрупувань. Це явище отримало назву поліклімаксу.
B.C. Ткаченко та А .П. Генов (1992) для заповідника "Кам'яні могили" Донецької області описали цікавий сукцесійний ряд степової екосистеми. Тут послідовно змінюють одна одну піонерна, типчакова, ковильова, корневищно-злакова, злаково-різнотравна, чагарникова та лісова фази. Примітно, що початкові фази цього ряду проявляють щорічні флуктуації, які сильно залежать від проникнення напівпаразитичної рослини дзвінець весняний. У поєднанні з риючою діяльністю сліпака це повертає степову екосистему до початкової фази. Розвиток відбувається мовби за своєрідними напівпетлями. Досить важливо, що в цьому випадку сукцесія є не тільки зміною характеру рослинного покриву, це динамічний процес усієї екосистеми.
2.9. Стійкість і саморегуляція екосистем
Лекція 3. Біосфера, її структура та закони функціонування
3.1. Поняття біосфери. Вчення В. І. Вернадського про біосферу
3.2. Структура біосфери
3.2.1. Основні типи речовини в біосфері
3.2.2. Атмосфера
3.2.3. Гідросфера
3.2.4. Літосфера
3.2.5. Живі організми в біосфері