До складу клітин еукаріотів входять поверхневий апарат, цитоплазма, ядро, немембранні та мембранні органели.
Поверхневий апарат клітин еукаріотів складається з:
o надмембранного комплексу (у рослин - клітинна стінка);
o плазматичної мембрани (плазмалеми);
o підмембранного комплексу (цитоскелету, що складається з мікрот-рубочок і мікрониток, і виконує опорну функцію).
Докладно зупинимось на особливостях будови, складу та функціях плазматичної мембрани, або плазмалеми, адже всі еукаріотичні клітини відокремлені нею від довкілля. Крім того, органели еукаріотичних клітини відділені мембранами від цитоплазми.
Плазматична мембрана, або плазмалема (від грецьк. plazma - виліплене, утворене, lema - оболонка), обмежує внутрішнє середовище клітини і виконує різноманітні функції: бар' єрну, обмін речовин, сприймає подразнення, забезпечує контакти між клітинами багатоклітинних організмів.
До складу клітинних мембран входять ліпіди (фосфоліпіди) (бар'єрні функції), білки (транспортні функції) та вуглеводи (рецепторні функції). Усі біологічні мембрани складаються з подвійного шару молекул фосфоліпідів, в який занурені молекули білків. Вуглеводи мембран зв' язані з білками та ліпідами.
Функції клітинних мембран:
o бар 'єрна - ізолюють клітину або її частину і підтримують в них сталість внутрішнього середовища (забезпечують ліпіди);
o транспортна - пропускають необхідні речовини всередину і назовні (забезпечують білки);
o рецепторна - забезпечують прийом інформації, реагують на зміни довкілля (забезпечують вуглеводи).
Плазмалема оточує клітину з усіх боків. Як же через неї переміщуються речовини? Обмін речовин між клітинами та міжклітинною речовиною забезпечується кількома способами транспорту: дифузія, полегшена дифузія (пасивний транспорт), активний транспорт, цитоз (екзо-цитоз, ендоцитоз).
Дифузія (від лат. diffusio - поширення) - переміщення молекул однієї речовини в іншій за їх безпосереднього контакту або крізь пори мембран, обумовлене тепловим рухом молекул. Цей процес (наприклад, переміщення таких речовин, як О2, СО2) не потребує витрат енергії.
Полегшена дифузія (пасивний транспорт) - це такий спосіб транспорту речовин через плазматичну мембрану клітини, який здійснюється білками-переносниками у напрямку меншої концентрації цих речовин. За такого способу проходять через мембрану окремі малі органічні молекули - глюкоза, деякі амінокислоти тощо.
Активний транспорт - це такий спосіб транспорту речовин через плазматичну мембрану клітини, пов' язаний із витратами енергії, оскільки не залежить від концентрації речовин, які мають потрапити в клітину або вийти з неї. За такого способу проходять йони та великі молекули, для яких мембрани є непроникними. На цей процес впливає різниця концентрацій іонів калію і натрію у зовнішньому середовищі та всередині клітини. Концентрація йонів калію всередині клітини вища, ніж ззовні, а йонів натрію - навпаки. Завдяки цьому йони натрію пересуваються в клітину, а калію - з неї. Але концентрація цих іонів у живій клітині і поза нею ніколи не вирівнюється, оскільки існує особливий механізм калій-натрієвого насосу, який іони натрію "відкачує" з клітини, а йони калію "закачує" в неї.
Цитоз - це спосіб транспорту великих молекул до клітини або з неї у мембранній упаковці. Цей процес властивий лише плазматичній мембрані. Розрізняють екзоцитоз (від грецьк. еІС20 - зовні, kutos - клітина) - мембранний транспорт з клітини та ендоцитоз (від грецьк. endon - внутрішній, - клітина) - мембранний транспорт у клітину.
Ендоцитоз - це такий спосіб надходження великих молекул або їхніх комплексів (наприклад, бактерії, віруси), коли вони не можуть пройти крізь мембрану, а надходять до клітини в мембранній упаковці. Розрізняють два основні види ендоцитозу: фагоцитоз і піноцитоз.
Фагоцитоз (від грецьк. phagos - пожирати, поглинати, kutos - клітина) - це активне захоплення твердих об'єктів - частинок органічних сполук, дрібних клітин та ін.
Піноцитоз (від грецьк. pino - п'ю, kutos - клітина) - це процес поглинання клітиною рідини разом із розчиненими у ній сполуками. Цей процес нагадує фагоцитоз, але відбувається здебільшого за рахунок вгинання мембрани.
Вода надходить до клітини та виходить з неї через плазмалему і ва-куолярну мембрану завдяки осмосу. Осмос - це явище, при якому відбувається вирівнювання концентрацій двох розчинів, які розділені напівпроникною мембраною за рахунок переміщення молекул розчинника з менш концентрованого до більш концентрованого розчину. Якщо концентрація солей у зовнішньому середовищі вища, ніж у клітині, то вода надходитиме з клітини у довкілля. Об'єм вакуолі і частково цитоплазми при цьому зменшиться, і цитоплазма почне відставати від клітинних стінок. Поступово вона може повністю відійти від стінок клітини і набути форми кулі. Це явище називається плазмолізом. Плазмоліз (від грецьк. plazma - оформлене, lyzis - розчинення) - явище відшарування цитоплазми з включеннями від клітинної стінки. Якщо таку плазмолізовану клітину помістити у дистильовану воду, то вода з навколишнього гіпотонічного середовища буде дифундувати до клітини, відновлюючи її первинний об'єм. Це явище називається деплазмолізом.
Цитоплазма (від грецьк. kytos - клітина, plazma - виліплене, утворене) - неоднорідний колоїдний вміст клітини. Цитоплазма еука-ріотичних клітин поділена мембранами на окремі функціональні ділянки, що забезпечує одночасний перебіг багатьох несумісних біохімічних процесів. До складу цитоплазми еукаріотичних клітин входять:
o цитозоль - місце, де відбуваються основні події в житті клітини. У клітині відбувається біосинтез (від грецьк. bios - життя, synthesis - сполука) - процес утворення органічних речовин. Проте біосинтез білків має виняткове значення. Від того, які білки синтезуються у клітині, залежить, які в ній будуть синтезуватися ліпіди, вуглеводи та інші речовини, оскільки для синтезу певної речовини потрібен відповідний фермент (білкової природи). Забезпечує зв'язок між усіма компонентами клітини;
o цитоскелет (від грецьк. kytos - клітина та skeleton ^ma) - висохле (тіло)) - опорно-рухова система клітини, що складається з мік-ротрубочок і мікрониток (забезпечує упорядковане розташування орга-нел, а також рухливість структур клітини);
o немембранні та мембранні органели (від грецьк. organon - орган, знаряддя, інструмент) - постійні структурні компоненти клітини, що виконують життєво необхідні функції;
o включення - непостійні структури, які можуть з'являтись у процесі життєдіяльності клітини, зникати і знову утворюватись; це запасні сполуки чи кінцеві продукти обміну речовин у вигляді краплин (жири), зерен (крохмаль, глікоген), кристалів (солі), пігментів (барвники) тощо.
Розглянемо докладно немембранні та мембранні органели. Немембранні органели
o Клітинний центр - немембранна органела, характерна для більшості еукаріот (рис.5). Клітинний центр завжди знаходиться в навко-лоядерній зоні, є геометричним центром клітини.
Клітинний центр складається з двох гранул - центріолей і мікрот-рубочок, що відходять від нього. Центріолі беруть участь у формуванні веретена поділу. При цьому вони розходяться до полюсів клітини і між ними натягуються нитки з мікротрубочок.
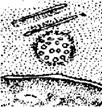
Рис. 5. Клітинний центр біля ядра
Після поділу материнської клітини в кожну з дочірніх потрапляє по одній центріолі. В період між двома поділами клітини ці структури подвоюються.
Функції клітинного центру ще остаточно не з'ясовані. Однак є підстави вважати, що він бере участь у формуванні мікротрубочок цитоплазми (тобто є головним організатором цитоскелета), веретена поділу клітини, джгутиків і війок.
У клітинах голонасінних і покритонасінних рослин клітинного центру немає, але є аналогічна органела.
o Рибосоми (від рибоза і грецьк. soma - тіло) - невеликі сферичні тільця, які лежать вільно або на мембранах ендоплазматичної сітки. До складу рибосом входить білок і рибосомальна РНК, магній. Ці органели клітини здійснюють біосинтез білків, властивих певному організмові (рис.6).

Рис. 6. Рибосоми на ЕПС (показані стрілкою)
Одномембранні органели
o Комплекс (апарат) Гольджі - система плоских цистерн, обмежених гладенькими мембранами (рис.7). Поруч із цистернами розташовані пухирці.

Рис. 7. Схема будови комплексу Гольджі
Функції комплексу Гольджі різноманітні:
а) забезпечення дозрівання, розподілу і транспорту синтезованих у клітині речовин;
б) бере участь в утворенні лізосом (ферменти, які входять до складу лізосом, синтезуються на мембранах зернистої ЕПС);
в) формування скоротливих вакуолей водоростей;
г) синтез полісахаридів для клітинної стінки.
o Ендоплазматична сітка (ЕПС), або ретикулум (від грецьк. endon - внутрішній, plazma - оформлене) - система мембран, що утворюють велику кількість каналів, трубочок, цистерн, завдяки чому значно збільшується внутрішня поверхня клітини і поділяється клітина на велику кількість комірок, що відіграє важливу роль у регуляції внутрішньоклітинних ферментних систем, транспорті речовин та перебігу процесів обміну.
У клітинах розрізняють два типи ЕПС:
1) гранулярна (шорстка, зерниста) - мембрани містять багато рибосом, які беруть участь у синтезі речовин білкової природи (рис.8); бере участь у синтезі мембранних ліпідів, ферментів лізосом;
2) гладенька (агранулярна, незерниста) - на мембранах рибосом немає; функції пов' язані з вуглеводним і жировим обміном, а також із знешкодженням отруйних для організму сполук.

Рис. 8. Гранулярна ЕПС
Таким чином, ЕПС бере участь у метаболізмі речовин, виконуючи роль внутрішньоклітинної, регуляторної і транспортної системи.
o Лізосоми (від грецьк. lyzis - розчинення, soma - тіло) - органели (рис.9), які містять різноманітні гідролітичні ферменти (гідролази), здатні розщеплювати органічні сполуки (білки, нуклеїнові кислоти, полісахариди, ліпіди), що надходять у клітину. Ферменти лізосом синтезуються в гранулярній ЕПС.
Лізосоми забезпечують процеси внутрішньоклітинного травлення.
У лізосомах перетравлюються мікроорганізми і віруси, окремі компоненти клітин, цілі клітини або їхні групи. Так знищуються дефектні органели, ушкоджені чи мертві клітини.

Рис. 9. Схема будови лізосоми
Лізосоми руйнують старі органели самої клітини, отже, беруть участь в самооновленні клітин. При голодуванні лізосоми перетравлюють запасні речовини та зайві органели, підтримуючи існування організму.
o Вакуолі (від лат. vacuus - порожній) - порожнини у цитоплазмі, заповнені рідиною і оточені мембраною:
1) вакуолі клітин рослин (рис.10) - утворюються з пухирців, які відокремлюються від ендоплазматичної сітки; заповнені клітинним соком - водним розчином органічних і неорганічних сполук, серед яких є продукти обміну речовин або пігменти; функції їх: підтримання тургору (від лат. turgere - бути набухлим), забезпечуючи пружність клітин і збереження їхньої форми; накопичення продуктів обміну речовин або запасних поживних речовин;
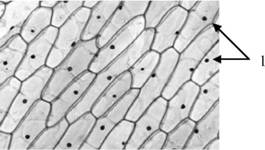
Рис.10. Вакуолі клітин рослин:
1 - вакуолі.
2) скоротливі вакуолі водоростей - формуються з елементів комплексу Гольджі; функції їх: регулювання внутрішньоклітинного тиску, виводячи надлишок води з клітини; беруть участь у виведенні деяких розчинних продуктів обміну речовин.
Двомембранні органели
o Пластиди - двомембранні органели, властиві тільки рослинній клітині. Вони утворюються з пропластид - ініціальних тілець меристем. Пропластиди - недиференційовані пластиди, що зустрічаються в меристемних клітинах коренів і пагонів, є попередниками пластид. Якщо світла немає, то в пропластиді з' являються проламелярні тільця (на-півкристалічні утворення в пластидах, розвиток яких зупинився через відсутність світла) і такі пластиди називають етіопластами. На світлі вони легко перетворюються на зелені пластиди - хлоропласти. Крім хлоропластів, розрізняють хромопласти, лейкопласти, амілопласти.
Розглянемо докладніше типи пластид:
1. Хлоропласти - зелені пластиди, в яких відбувається фотосинтез. Вони зустрічаються переважно в клітинах паренхіми, їх немає в меристемі, кількість їх в клітині від одного до сотень, діаметр 5-8 мкм, товщина до 1 мкм. Хлоропласти мають округлу форму. У них є дві мембрани - зовнішня та внутрішня. Внутрішня мембрана утворює направлені всередину сплощені вирости - ламели, які занурені в гідрофільний білковий матрикс, або строму. Основна структурна одиниця хлоропластів - тилакоїд - плоский диск, оточений одинарною мембраною. Тилакоїди зібрані в щось подібне до купки монет, які називають гранами (від лат. granum - зерно, крупинка). Саме в мембранах гран знаходиться зелений пігмент - хлорофіл, де відбувається фотосинтез. У гранах виявлено перфорації (від лат. рег/огаЫо - просвердлювання), крізь які мембрани гран поєднуються, а значить і їх внутріш-ньотилакоїдний простір сполучається за допомогою вузеньких трубочок фрет. Тилакоїди, які зв' язують між собою грани, називають тилакоїдами строми.
Чому хлорофіл зелений? Колір речовини залежить від того, які промені світла вона відбиває, а які - поглинає. Хлорофіл, основний пігмент фотосинтезу, відбиває зелені промені, а поглинає фіолетові, червоні й сині.
У матриксі хлоропластів знаходяться також і інші структури: ДНК (замкнена в кільце), іРНК, тРНК, рибосоми, зерна крохмалю. Отже, хлоропласти мають власну спадкову інформацію та білоксинтезуючу систему. Для пояснення цих фактів на початку ХХ ст. було запропоновано ендосимбіотичну гіпотезу, за якою хлоропласти - нащадки про-каріотичних клітин, що перейшли до життя всередині інших клітин. У хлоропластах також синтезуються деякі ліпіди, білки мембран тилакої-дів, ферменти, які каталізують реакції фотосинтезу. Крім того, в них, як і в мітохондріях, синтезується АТФ.
2. Хромопласти - жовті, оранжеві або червоні пластиди, які за рахунок пігментів (переважно каротиноїдів) зумовлюють відповідне забарвлення пелюсток квіток, плодів, коренеплодів тощо. Хромопласти, як правило, розвиваються з хлоропластів, мають приблизно такі ж розміри та форму, досить схожі і за структурою. Однак замість системи фотосинтетичних мембран в них знаходяться структури, багаті на каротиноїди. За своєю внутрішньою структурою хромопласти поділяють на 5 типів:
1) глобулярний - характерний для більшості пелюсток квіток, плас-тоглобули з каротиноїдами діаметром до 150 нм;
2) мембранний - має до 25 різних типів концентричних мембран, теж зустрічається в пелюстках квіток;
3) трубчастий - характеризується наявністю волокон 15-80 нм, які містять білково-каротиноїдні комплекси;
4) ретикулотрубчастий - густа мережа розгалужених непаралель-них трубочок;
5) кристалічний - містить каротиноїди у формі кристалів (наприклад, у плодах помідорів, де каротин - лікопін, знаходиться в кристалічних трубочках довжиною 15-48 мкм).
Фізіологічна функція хромопластів невідома. Яскраве забарвлення, можливо, принаджує комах для запилення та поширення насіння.
3. Лейкопласти - не пігментовані пластиди. Оболонка лейкопласта складається з двох мембран: зовнішньої і внутрішньої. Внутрішня мембрана вростає в строму, утворює малочисельні тилакоїди. У стромі є ДНК, рибосоми, а також ферменти, які здійснюють синтез і гідроліз запасаючих речовин. Лейкопласти можуть перетворюватися в хлоропласти, рідше у хромопласти.
4. Амілопласти - це зрілі пластиди, вміст яких складається майже повністю з крохмальних зерен. Зустрічаються в запасних тканинах, а саме: сім'ядолі, ендоспермі, бульбі, а також у кореневому чохлику. Крохмальні зерна знаходяться в матриксі (стромі), яка містить також ДНК та кілька рибосом.
Функція амілопластів - синтезувати крохмаль із цукрози, яка надходить сюди з фотосинтезуючих органів, а також запасати його з наступними витратами у разі потреби, наприклад, при проростанні. В кореневому чохлику вони виконують зовсім іншу роль, пов' язану з сприйняттям гравітації.
Усі пластиди здатні до взаємоперетворення. Етіопласти на світлі перетворюються на хлоропласти. Амілопласти утворюються як проміжні форми на шляху розвитку етіопластів та хлоропластів. Так, навіть в таких типових амілопластах бульб картоплі на світлі розвивається тила-коїдна система (позеленіння бульб). Обернений цьому процес спостерігається в апельсинах та коренеплодах моркви. Осіннє пожовтіння листя пов' язано з перетворенням хлоропластів на хромопласти.
Речовини, що надають певного кольору пластидам і клітинному соку - пігменти. Пігментами називають сполуки, які вибірково поглинають світло у видимій частині спектра. Пігменти пластид належать до трьох класів: -хлорофілів, каротиноїдів, фікобілінів.
Усі ці пігменти локалізовані в особливих частинах клітини - фіко-білісомах, які певним чином формують впорядковані ансамблі на поверхні тилакоїдних мембран. Як правило, в більшості водоростей фікобі-ліни присутні в значно більшій концентрації порівняно з хлорофілами, тому саме вони і визначають їх забарвлення. Між собою вони також зустрічаються в різних співвідношеннях, причому залежно від умов освітлення формується переважно такий набір пігментів, який найкраще використовує відповідний спектр. Таке явище хроматичної адаптації має важливе адаптивне значення.
o Мітохондрії (від грецьк. mitos - нитка, chondrion - зернятко) - органели двомембранної будови, основна функція яких полягає у виробленні майже всієї енергії клітини (синтез АТФ) (рис.11). Це відбувається шляхом поступового окиснення органічних сполук, які поступають до клітини. Звільнена при цьому енергія використовується мітохондріями для синтезу молекул АТФ і АДФ, які можуть депонуватися в мітохондріях і використовуватися за потребою.
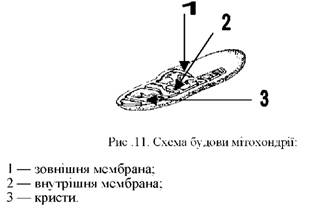
Поверхневий апарат мітохондрій складається з двох мембран (рис.11). Зовнішня мембрана гладенька, вона відмежовує цю органелу від цитоплазми. Внутрішня мембрана утворює випинання всередину мітохондрій у вигляді трубчастих чи гребінчастих утворів - крист. На поверхні внутрішньої мембрани, оберненій всередину мітохондрії, є особливі утвори, які містять комплекс ферментів, потрібних для синтезу АТФ. Внутрішній простір мітохондрій заповнений напіврідкою речовиною - матриксом. У ньому містяться кільцева молекула ДНК, іРНК, тРНК, рибосоми, тобто мітохондрії, подібно до хлоропластів, мають власну спадкову інформацію та білоксинтезуючу систему. За ен-досимбіотичною гіпотезою мітохондрії, як і хлоропласти, - нащадки прокаріотичних клітин, що перейшли до життя всередині інших клітин.
o Ядро (від грецьк. karion - ядро; від лат. nucleus - ядро) - складова частина живої клітини, яка зберігає спадкову інформацію, передає її дочірнім клітинам під час поділу і керує життєвими процесами. До складу ядра входять (рис.12):
■ поверхневий апарат ядра - ядерна оболонка (складається з двох мембран - зовнішньої та внутрішньої) з порами (місця, де зовнішня і внутрішня мембрани з'єднуються; кожна пора закрита поросомою, яка регулює транспорт речовин крізь пору), білкова ядерна пластинка (стійкий опорний елемент ядра, що прилягає до внутрішньої мембрани зсередини; надає йому форми і служить для прикріплення хромосом);
■ каріоплазма (від грецьк. karion - ядро, plazma - оформлене), або ядерний сік - прозоре напіврідке внутрішнє середовище ядра, оточене ядерною оболонкою, в якому проходять усі реакції; у каріопла-змі розташовані комплекси нуклеїнових кислот з білками (це - хроматин: ДНК+білки тарибонуклеопротеїдні комплекси: РНК+білки);
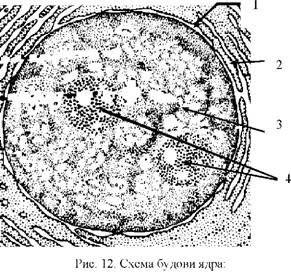
1 - ядерна оболонка;
2 - пори;
3 - каріоплазма;
4 - ядерця.
■ хроматин (ДНК+білки) - ниткоподібні структури, з яких під час поділу клітини формуються хромосоми; сукупність ознак хромосомного набору (кількість хромосом, їхня форма і розміри) - це каріотип;
■ ядерце, або рибонуклеопротеїдні комплекси (РНК+білки) - утворюється на тих ділянках хромосом, де відбувається синтез рРНК, містить гранули - попередники рибосом;
■ ядерний матрикс - ниткоподібні опорні структури, які забезпечують упорядковане розташування хромосом, а також сполучають між собою ядерця, нитки хроматину, ядерні пори тощо.
Є клітини (наприклад, ситоподібні трубки рослин), у яких на певному етапі розвитку ядро зникає. Внаслідок цього більшу частину життя такі клітини позбавлені ядра, тому вони не здатні до розмноження і швидко гинуть. Функції ядра:
o збереження спадкової інформації, закодованої в ДНК;
o забезпечує реалізацію спадкової інформації завдяки транскрипції (синтезу іРНК); цей процес відбувається в ядрі на деконденсованих (розтягнених) ділянках хромосом. При фарбуванні клітини лужними барвниками ці ділянки не забарвлюються і тому непомітні під мікроскопом (конденсовані, або скручені, ділянки забарвлюються і набувають вигляду темних гранул). Місця, де вони розташовані, виглядають прозорими (світлими) зонами. Отже, якщо в ядрі багато світлих зон, можна передбачити інтенсивну транскрипцію;
o забезпечує передачу спадкової інформації від материнської клітини дочірнім;
o синтезує тРНК, рРНК;
o забезпечує формування рибосом (за участю ядерець).
o Клітинний цикл - період існування клітини від початку останнього поділу до наступного або ж від початку останнього поділу клітини до її загибелі. Клітинний цикл за оптимальних умов триває у клітин еукаріотів - 10-80 годин і більше (для порівняння: у бактерій - 20-30 хвилин).
Клітинний цикл складається з періоду поділу клітини і проміжку між двома поділами - інтерфази. Інтерфаза (від лат. inter - між, посеред і грецьк. phasis - поява) - частина клітинного циклу між двома послідовними поділами клітини.
В інтерфазі виділяють три періоди, які складають до 90% часу всього клітинного циклу: передсинтетичний (GЎ), синтетичний (S), пост-синтетичний (G2).
Передсинтетичний період (G1) іде безпосередньо за поділом клітини. У цей період нагромаджуються РНК і білок, які необхідні для утворення клітинних структур та росту клітини. Це найтриваліший період (може тривати від 10 годин до кількох діб).
Синтетичний період (S) характеризується інтенсивним синтезом молекул ядерної ДНК, які утворюють хромосоми. Кожна хромосома - це пара однакових за генетичним матеріалом хроматид, з'єднаних між собою центромерою. На ДНК-матрицях синтезуються копії ДНК: кожний з двох полінуклеотидних ланцюгів ДНК слугує матрицею (шаблоном) для синтезу комплементарного йому другого ланцюга. У результаті цього відбувається подвоєння молекул ДНК й утворюються дві однакові молекули. Цей процес називають реплікацією (англ. replication - копіювання, від лат. replicatio - відбиття), або редуплікацією (від лат. reduplico - подвоюю). У цей період самоподвоюється клітинний центр, внаслідок чого утворюються дві центріолі. Синтез РНК і білка продовжується. Тривалість синтетичного періоду - 6-10 годин.
Постсинтетичний період (G2): відбувається нагромадження енергії у формі АТФ за рахунок інтенсивного функціонування мітохондрій.
Ця енергія необхідна для поділу клітини. В цей період триває синтез РНК і білків, переважно ядерних. Тривалість постсинтетичного періоду 3-4 години.
Процес поділу еукаріотичних клітин супроводжується утворенням особливого веретена поділу, що забезпечує розподіл спадкового матеріалу між дочірніми клітинами.
Еукаріотичним клітинам властивий поділ шляхом мітозу, мейозу, амітозу.
Мітоз (непрямий поділ) (від грецьк. mitos - нитка) - поділ ядра, що забезпечує рівний розподіл генетичного матеріалу між дочірніми клітинами і спадкоємність хромосом у ряду клітинних поколінь. Внаслідок мітозу з однієї материнської клітини утворюються дві дочірні.
У процесі мітозу послідовно відбуваються фази: профаза, метафаза, анафаза, телофаза.
Мітоз починається з профази (від грецьк. pro - до, phasis - поява), на початку якої розбирається оболонка ядра на окремі мікропухирці (рис. 13). Хромосоми втрачають зв'язок з ядерною оболонкою і починають конденсуватися (ущільнюватися), завдяки цьому їх можна побачити у світловий мікроскоп.

На початку профази кожна хромосома представлена одиничною структурою, всередині - кожна хромосома вже складається з двох ідентичних хроматид (майбутніх дочірніх хромосом), що утворилися при реплікації ДНК в період інтерфази. Між хроматидами з'являється щілиноподібний простір, який весь час розширюється. Зникає ядерце, розбираються мікротрубочки цитоскелета, комплекс Гольджі та ЕПС розпадаються на мікроскопічні пухирці. У кінці профази центріолі клітинного центру парами розходяться до полюсів клітини і стимулюють утворення мікротрубочок веретена поділу.
У метафазі (від грецьк. meta - після, phasis - поява) хромосоми знаходяться в упорядкованому стані у ділянці екватора клітини і прикріплюються до мікротрубочок веретена поділу (рис. 14). Хроматиди відштовхуються одна від одної і залишаються з' єднаними тільки в зоні первинної перетяжки.
Рис. 14. Схема метафази
В анафазі (від грецьк. ana - навпроти, phasis - поява) хроматиди (дочірні хромосоми) відокремлюються і розходяться до полюсів материнської клітини. Нитки веретена скорочуються і розтягують хромосоми до полюсів клітини (рис. 15).

Рис. 15. Схема анафази
У телофазі (від грецьк. telos - кінець, phasis - поява) (рис.16) хромосоми перестають рухатись, розміщуються на протилежних полюсах клітини, деконденсуються, або деспіралізуються (набувають розтягнутого стану), втрачають свою індивідуальність, набухають, зливаються у загальний клубок. Навколо них починає утворюватись нова ядерна оболонка.

Рис. 16. Схема телофази
У телофазі закінчується процес руйнування апарату ділення. Головним моментом телофази є поділ цитоплазми - цитокінез. При цьому до кожної дочірньої клітини потрапляє приблизно рівна кількість органел.
Мітоз закінчується утворенням двох дочірніх клітин. Загальна схема мітозу подана на рис. 17.
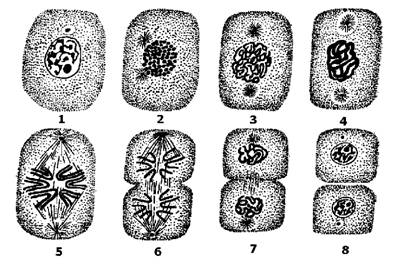
Рис. 17. Послідовні стадії мітозу:
1 - вихідна (материнська) клітина;
2 - 4 - профаза;
5 - метафаза;
6 - анафаза;
7 - телофаза;
8 - дві дочірні клітини.
Біологічне значення мітозу: забезпечує точну передачу спадкової інформації впродовж низки послідовних клітинних циклів, стабільність каріотипу організмів певного виду.
Крім того, завдяки мітозу відбувається регенерація тканин та органів. Регенерація (від лат. regeneratio - відновлення) - відновлення організмом утрачених чи пошкоджених органів або тканин.
Дочірні клітини, які утворилися внаслідок мітозу, відрізняються від материнської: вони удвічі менші за материнську. Ріст клітин та виконання ними специфічних функцій відбувається в інтерфазі.
Чи обмежена можлива кількість поділів клітини? На це питання немає однозначної відповіді.
Які механізми можуть обмежувати число клітинних циклів у нащадків якоїсь клітини? Нині найвірогіднішою вважається гіпотеза, згідно з якою термін життя клітини залежить від довжини певних ділянок хромосом. На кінцях кожної хромосоми є ділянки - теломери, які з кожним поділом зменшуються. Це своєрідний біологічний годинник життя клітини. Таким чином, клітини, утворені при мітозі, дещо відрізняються від материнських.
Мейоз (від грецьк. meiosis - зменшення) - поділ ядра, при якому відбувається зменшення кількості хромосом удвічі, причому з однієї диплоїдної (2п) клітини утворюються чотири гаплоїдні (п). Мейоз здійснюється тільки у процесі утворення статевих клітин. Під час мейозу, на відміну від мітозу, відбуваються два послідовні поділи, інтерфаза між якими вкорочена, а у клітин рослин її взагалі немає. Кожний з цих поділів, як і мітоз, має чотири послідовні фази: профазу, метафазу, анафазу, телофазу. В результаті двох послідовних мейотичних поділів з однієї клітини з диплоїдним набором хромосом утворюється чотири клітини з гаплоїдним набором хромосом.
Біологічне значення мейозу. Мейоз являє собою досконалий механізм, який забезпечує сталість каріотипу видів, які розмножуються статевим способом. Завдяки двом поділам статеві клітини мають половинний порівняно з нестатевими набір хромосом. А набір хромосом, характерний для організмів певного виду, відновлюється під час запліднення. В результаті утворюється зигота (від грецьк. zygotos - з' єднання докупи) - диплоїдна клітина, що утворюється внаслідок злиття чоловічої й жіночої статевих клітин (гамет).
Мейоз також забезпечує і спадкову мінливість організмів. Тому клітини, що утворилися внаслідок мейозу, можуть мати відмінний від материнської набір спадкової інформації.
Амітоз - прямий поділ клітини. На відміну від мітозу при амітозі зберігається інтерфазна структура ядра і хромосоми під оптичним мікроскопом не видимі. Ядро при цьому ділиться шляхом перетяжки на дві відносно рівні частини. Точного розподілу ДНК між ними не буває. Інколи після поділу ядра перешнуровується і цитоплазма і утворюються дві клітини. В інших випадках клітина залишається двоядерною. Клітини, що утворилися внаслідок амітозу, мають порушений набір хромосом і, як правило, швидко гинуть.
Питання для самоперевірки
ЛЕКЦІЯ 5. Тканинний рівень організації живої матерії. Рослинні тканини
1. Поняття про тканини
2. Рослинні тканини
Питання для самоперевірки
ЛЕКЦІЯ 6. Органографія. вегетативні органи рослинного організму. Корінь
1. Органографія
2. Вегетативні органи рослинного організму
2.1. Корінь