Розміри популяції можуть зростати в результаті імміграції (додаються особини ззовні) або за рахунок розмноження особин. На зміни в чисельному складі популяції суттєвий вплив мають кліматичні умови, які висвітлено в попередньому розділі (екофактори — температура, вологість і т.д.). Нерідко лімітуючим фактором, як уже доведено, виступають вороги, їжа тощо. Коливання чисельності відбуваються циклічно, їх можна назвати циклами. Але дослідження таких циклів потребує тривалого часу і залежить від періоду між максимумом та мінімумом чисельності даної популяції. З огляду на період настання статевої зрілості, вагітність, у кожного виду ці параметри різні. У маленької тваринки, такої, як бурозубка, ці періоди набагато коротші, ніж у таких, як копитні, слони. Тобто, щоб прослідкувати цей процес, екологу необхідно володіти інформацією за той відтинок часу, за який проходить численна зміна генерацій (поколінь), і знати умови існування цієї популяції. Набагато простіше цю інформацію можна здобути в лабораторних умовах, де в ході експерименту дослідник інколи штучно, а деколи й підсвідомо створює сприятливі умови існування (пацюк, дрозофіла і т.д.).
Коливання чисельності популяції графічно можна зобразити у формі синусоїди (рис. 3.4), для побудови якої необхідно проводити дослідження протягом тривалого часу. Ця синусоїда складається із фрагментів" що можуть відхилятись від ідеальної кривої. Важливим моментом є той факт, що коливний процес відбувається навколо уявної лінії, яка і буде ідеальним графічним вираженням чисельності популяції. Також слід відзначити: коливання чисельності особин популяції можливе в певних межах, отож цілком слушно тут фігурує поняття мінімальної чисельності популяції. Якщо чисельність особин сягає покажчиків, нижчих від мінімальної чисельності, то вона зникає.
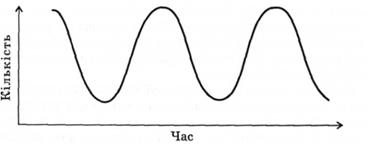
Рис. 3.4. Циклічне коливання чисельності популяції
Розміри популяції не можуть бути постійними через зміни плодючості, смертності, а нерідко і того, й іншого. При вивченні розмірів популяцій та їх змін завжди намагаються встановити ключовий фактор — той, який відповідає за найбільшу частину змін, що відбуваються при зміні поколінь. Як правило, цей ключовий фактор впливає на смертність.
Доведено, що коливання розмірів популяції проходить не хаотично. Насправді є низка факторів, які втримують стан популяції* в певних межах. Це фактори, які знижують чисельність і сприяють смертності та найкраще діють при збільшенні щільності. Такими факторами можуть бути нестача їжі, збільшення кількості ворогів тощо.
3.6. Зростання чисельності популяції, криві вростання та виживання
Якщо народжуваність у популяції перевищуватиме смертність, така популяція буде збільшуватись. Яскравим прикладом такого явища є зростання народонаселення Землі. Підраховано, що тільки упродовж XX ст. народонаселення зросло більш ніж удвічі. Тобто внаслідок якісного стрибка людства, науково-технічного прогресу людство створило певні умови, які і спричинили таке різке зростання.
Загальний перебіг зміни чисельності особин у популяції визначається рівнянням: Nt+1 = N+B-D+I-E, де N — чисельність особин у популяції, В — народжуваність, D — смертність, І — імміґрація, Е — еміґрація, t — час.
Розміри популяції можуть збільшуватися за рахунок або великої народжуваності, або високої імміграції, або поєднання обох цих факторів. Знижують розмір популяції смертність та еміґрація особин за її межі.
Щоб ясно уявити закономірності збільшення популяції, доцільно розглянути модель зростання поігуляції дріжджів, які потрапили на свіжу культурну речовину (рис. 3.5). В такому новому та сприятливому середовищі умови для збільшення популяції оптимальні, отож невдовзі спостерігатиметься експоненціальне зростання популяції. У свіжому поживному середовищі зростання йтиме поступово, досягаючи максимальної чисельності. Затримка у збільшенні популяції на початкових етапах пов'язана із адаптацією до нових умов середовища. Намальована нами крива — це експоненціальна, або логарифмічна, крива. В наступних етапах життєдіяльності популяції настає період, коли експоненціальний розвиток неможливий. Таке може статися з різних причин — зменшення ресурсів
живлення, накопичення продуктів метаболізму тощо. Як наслідок — процес зростання популяції поступово уповільнюється і крива зростання набуває s-подібну форму.
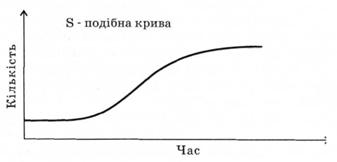
Рис. 3.5. Модель зростання чисельності популяції дріжджів
Існує й інший тип зростання чисельності популяції, коли експоненціальне зростання триває до раптового зменшення кількості організмів (рис. 3.6). Це явище може статись за рахунок різкого зменшення ресурсу, території тощо. Такий тип кривої зростання дістав назву J-подібної кривої. Слід відзначити, що в обох випадках експоненціальне зростання простежується на початкових етапах росту.
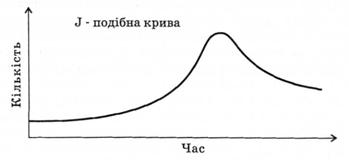
Рис. 3.6. Модель зростання чисельності популяції
Отже, ми розглянули дві моделі зростання популяції. Водночас слід зауважити, що побудова таких кривих можлива тільки за умови більш-менш стабільного існування екосистеми. Тобто там, де чинники системи не діють як лімітуючі на зростання популяції.
Щойно змальовані моделі в чистому виді, як правило, в природі не існують. Якщо деякі подібності можна зустріти в природі при розселенні і освоєнні видом нових територій (це наочно можна проілюструвати розселенням горлиці кільчастої в Центральній Європі), то на територіях, де види-інтродуценти вже ввійшли до складу екосистем, такого спостерігатись не буде. Однак такі моделі дають нам змогу розуміти закономірності зростання чисельності популяцій, прогнозувати поведінку виду в нових умовах, управляти і корегувати чисельність "корисних" і "шкідливих" видів.
Одним із основних факторів, які впливають на розміри популяції, є процент особин, які гинуть до досягнення ними статевої зрілості. Аби чисельність популяції залишалась сталою, в середньому тільки два потомки кожної пари мають доживати до репродуктивного віку. Щоб дістати криву виживання, доцільно почати з певної популяції новонароджених особин і потім відмічати кількість особин, що вижили, залежно від часу. Накреслюючи криві виживання для окремих видів, можна визначати смертність для особин різного віку і таким чином вияснити, в якому віці цей вид найбільш вразливий. Якщо встановити причини смерті, можна зрозуміти, як регулюється величина популяції.
Криву виживання можна дістати, якщо розпочати спостереження за певною популяцією, відстежуючи тільки новонароджених особин, і відмічати число або відсоток особин, які залишаються жити, залежно від часу. Більшість тварин і рослин старіють, що виявляється передусім у зменшенні кількості особин після досягнення репродуктивного періоду (рис. 3.7).
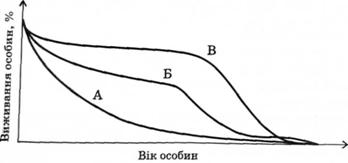
Причинами цього явища є чимало факторів, але, як правило, в пострепродуктивний період організм поступово втрачає свою захисну здатність. Крива А — характерна для видів, де смертність є більш-менш сталою одиницею у всі періоди розвитку. Для більшості безхребетних така крива е типовою. Крива Б — характерна для популяцій організмів з високими показниками смертності в ранній дорепродуктивний період. Така крива характерна для муфлонів, гірських кіз. Крива В — близька до ідеальної кривої, оскільки пересвідчуємось, що смертність тривалий період часу поступається віку, а старіння є головним фактором смертності. За приклад можна взяти популяцію людей на нашій планеті. Велика кількість людей помирає внаслідок старіння, але середній вік не перевищує 75 років. Невелике відхилення на початкових фазах пов'язано із дитячою (дорепродуктивною) смертністю.
Завершуючи розгляд питань динаміки чисельності популяції, слід відзначити: процес коливання чисельності неперервний і може змінюватись в часі, як наслідок адаптаційних змін. Зникнення цього явища можливе тільки у зв'язку зі зникненням виду. Питання динаміки популяцій є основою для розуміння ширших питань, таких, як динаміка угруповання, екосистеми, біосфери в цілому.
3.7. Практичне значення вчення про популяції
Питання для самоперевірки
РОЗДІЛ 4 ЕКОЛОГІЯ УГРУПОВАНЬ
4.1. Біоценози (угруповання). їх таксономічний та функціональний склад. Систематика біоценозів
4.2. Трофічна, просторова, екологічна структури біоценозу
Трофічна структура біоценозу
Просторова структура угруповання
Екологічна структура біоценозу
4.3. Видова структура угруповань та способи її виявлення