Нервовий центр - це складна сукупність нейронів, необхідних для здійснення рефлекторного акту чи регуляції певної функції організму. Діяльність нервових центрів грунтується на взаємодії двох процесів збудження і гальмування.
Збудження - активний фізіологічний процес яким нервові клітини відповідають на зовнішню дію.
Збудженню характерні такі властивості:
- однобічне проведення збудження - це здатність проводити збудження в одному напрямку, що зумовлена властивістю хімічних синап-сів проводити збудження від аферентного аксона до еферентного нейрона через синапс;
- уповільнене проведення збудження в нервовому центрі зумовлене синаптичною затримкою, тобто часом, потрібним для повного розвитку дії медіатору на постсинаптичну мембрану проміжних та еферентних нейронів;
- сумація збуджень: часова сумація виникає тоді, коли допорогові подразнення багаторазово повторюються через короткі інтервали часу; просторова сумація виникає тоді, коли кілька допорогових подразників діють на різні ділянки рецептивного поля якогось рефлексу;
- трансформація ритму збудження: частота імпульсів, які нервові центри надсилають до виконавчих органів, певною мірою визначаються силою і частотою подразнення рецепторів;
- післядія збудження - це продовження рефлекторної відповіді після припинення подразнення. Післядія збудження нервового центру може зберігатися у вигляді слідів і після закінчення збудження. Тривалість збереження цих слідів різна - від кількох секунд до багатьох років, що має значення для процесів пам'яті;
- тонус нервових центрів - це стале незначне збудження нейронів нервового центру, у створенні і підтриманні якого беруть участь аферентні імпульси, що надходять від периферичних рецепторів ЦНС, а також різні гуморальні подразники;
- принцип домінанти: в ЦНС постійно виникають і можуть тривалий час існувати домінуючі осередки збудження, які підкорюють собі роботу інших нервових центрів; цей принцип був сформульований О.О. Ухтомським як загальний принцип функції нервових центрів;
Гальмування - активний нервовий процес, який веде до зменшення чи припинення збудження в локальній ділянці нервової тканини. На відміну від процесу збудження, гальмування може розвиватися тільки у вигляді локального процесу і тому завжди пов'язане з існуванням спеціальних гальмівних синапсів. Гальмівними нейронами у спинному мозку є вставні нейрони - клітини Ренто, в мозочку - грушоподібні нейрони (Пуркіньє). Розрізняють кілька видів гальмування у ЦНС:
- постсинаптичне гальмування полягає в тому, що медіатор, який виділяється пресинаптичним закінченням гальмівних нейронів, змінює властивості постсинаптичної мембрани таким чином, що вона не може генерувати збудження. Постсинаптичне гальмування може бути прямим і зворотним. Пряме постсинаптичне гальмування виникає у постсинаптичному нейроні внаслідок гіперполяризації його мембрани під час розвитку гальмівного постсинаптичного потенціалу під впливом потенціалу дії будь-якого гальмівного нейрона. Зворотне постсинаптичне гальмування здійснюється у нейтронному ланцюзі, який складається з рухового нейрона та вставного гальмівного нейрона (клітини Ренто). Цей гальмівний нейрон спричинює пригнічення розрядів рухового нейрона;
- пресинаптичне гальмування здійснюється за допомогою спеціальних вставних нейронів і значно триваліше за постсинаптичне. Воно особливо ефективне для обробки інформації, яка надходить до нейрона різними пресинаптичними шляхами, оскільки в цьому разі збудження, що надходить через один із входів, може бути вибірково зменшеним або навіть повністю пригнічене, якщо немає інших впливів на інші синоптичні ходи.
Структурною основою цього гальмування є аксоаксонні синапси, утворені закінченнями аксонів гальмівних нейронів на пресинаптичних нервових закінченнях збуджувальних нейшнів (рис. 27).
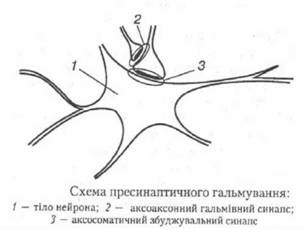
Рис. 27. (за Чайченко П. М., Цибенко В. О., Сокун В. Д., 2003)
Медіатор, що виділяється на цих синапсах (гамма-аміномасляна кислота), викликає деполяризацію нервових закінчень за рахунок збільшення проникності мембрани для іонів СІ-. Припускають, що пресинаптичне гальмування пригнічує сигнали, біологічне значення яких у певний момент є незначним, і пропускає інформацію, що є найважливішою для організму.
При постсинаптичному гальмуванні імпульс, який прийшов до гальмівного синапсу, зумовлює гіперполяризацію постсинаптичної мембрани. При цьому зростає величина мембранного потенціалу і виникає гальмівний постсинаптичний потенціал (ГПСП), який є локальною гіперполяризацією мембрани, що виникає внаслідок підвищення її проникності до СІ- або К . Постсинаптичне гальмування може бути прямим і зворотним.
Пряме постсинаптичне гальмування виникає у постсинаптичному нейроні внаслідок гіперполяризації його мембрани під час розвитку ГПСП під впливом ПД будь-якого гальмівного нейрона.
Зворотне постсинаптичне гальмування здійснюється у нейронному ланцюзі, який складається з рухового нейрона та вставного гальмівного нейрона (клітини Реншо). Імпульси від збудження рухового нейрона не тільки прямують через аксон, а й поширюються від нього, і активують вставний нейрон-клітину Реншо. Цей гальмівний нейрон спричинює пригнічення розрядів рухового нейрона. Отже, з цих двох нейронів утворюється контур з негативним зворотним зв'язком, який дає змогу стабілізувати частоту розрядів рухового нейрона, обмежуючи надмірну імпульсацію до ефекторного органа.
У нейронах кори головного мозку і таламусу виявлено два види постсинаптичного гальмування:
1) соматичне, яке виникає під час збудження гальмівних синапсів, розміщених на тілі і найближчих дендритах нейрона внаслідок генерації високоамплітудних ГПСП, що блокують реакції нейрона на будь-яку аферентну імпульсацію;
2) дендритне, яке з'являється під час збудження гальмівних аксо-дендритних і дендро-дендритних синапсів, розміщених на дистальних ділянках дендритів. ГПСП, що виникає при цьому, завдяки своїй невеликій амплітуді і значній відстані між місцем виникнення і аксонним горбком не здатні викликати значні зміни мембранного потенціалу нейрона. Отже, дендритне гальмування має локальний характер і забезпечує вибіркове пригнічення аферентного входу до нейрона через певні частини дендритного дерева, відіграючи важливу роль в інтегративній діяльності мозку (М.П.Сєрков).
Вторинне гальмування здійснюється без участі спеціальних гальмівних структур і розвивається в збудливих синапсах при дії подразників надмірної сили.
Численними дослідами встановлено, що збудження в одних центрах нервової системи викликає гальмування в інших. Наприклад, коли м'язи-згиначі правої ноги і розгиначі лівої скорочуються, розгиначі правої ноги і згиначі лівої перебувають у розслабленому стані. Така координована діяльність м'язів нижніх кінцівок пояснюється тим, що збудження в нервових центрах згиначів викликає гальмівний стан в центрах цієї ж кінцівки. При цьому збудження в центрі згиначів однієї кінцівки зв'язане з гальмуванням у центрі згиначів другої. Згідно з ученням видатного російського фізіолога М.Е.Введенського, гальмування і збудження є лише два прояви одного і того самого нервового процесу, які залежать лише від сили і частоти подразнення.
Периферичні нервові волокна, розплющені або частково перерізані, повільно регенерують, якщо тіло клітини і сегменти мієлінової оболонки неперервні. Регенерація волокон не відбувається в головному та спинному мозку. Ушкоджені нервові волокна трансформуються в тканину рубця та інактивуються.
Якщо нервове волокно ушкоджується в місці, найбільш віддаленому від тіла клітини, настає дегенерація внаслідок нестачі життєво важливих білків та ензимів, і мієлінова оболонка стає порожнистою.
Неушкоджене тіло нейрона стимулює ріст декількох нервових відростків у залишках волокна. Один з цих відростків може прорости через порожню, але неушкоджену мієлінову оболонку.
Нове нервове волокно, яке росте зі швидкістю близько 1,5 мм на день, досягає попереднього місця розташування, функція і відчуття повільно відновлюються, а невикористані нервові відростки дегенерують.
Значення процесів гальмування полягає в тому, що:
o разом зі збудженням процес гальмування відіграє значну роль у пристосуванні організму до навколишнього середовища;
o гальмування відіграє важливу роль у формуванні умовних рефлексів, звільняючи ЦНС від перероблення несуттєвої інформації.
Скупчення відростків нервових клітин - нервових волокон - вкрите зверху сполучнотканинною оболонкою і називається нервом. Кожен нерв складається з дуже багатьох нервових волокон. Нерви, по яких збудження поширюється в напрямку ЦНС, називаються доцентровими, або аферентними, або інформаційними. Якщо в складі нерва є нервові волокна, які передають збудження від центральної нервової системи (ЦНС) до іннервованого органа, то такі нерви називають відцентровими, або еферентними, або руховими. Більшість нервів змішані.
9. Основні етапи розвитку нервової системи
З історії науки
Еволюційний процес
ЛЕКЦІЯ 3. Будова, функції та розвиток спинного мозку
1. Будова спинного мозку
1.1. Оболонки спинного мозку
1.2. Лікворна система мозку
1.3. Поперечний переріз спинного мозку
1.4. Спинномозкові нервові сплетення