Квітка диференціюється з апікальної меристеми головної чи бічних осей після переходу її в репродуктивну фазу. Цей процес контролюється гормонально та стимулюється різноманітними зовнішніми факторами, у першу чергу зміною довжини світлового дня. Поблизу апікальної меристеми у вигляді примордіальних горбочків виникають зачатки частин квітки. Першими закладаються чашолистки, примордії пелюсток, потім послідовно тичинки та плодолистики, які зростаючись, утворюють маточку і до яких, як правило, прикріплені насінні зачатки, у яких формується жіночий гаметофіт.
Формування тичинки відбувається у такій послідовності. Спочатку виникає пиляк, а пізніше тичинкова нитка. Через неї і в'язальце проходить провідний пучок. У пиляках формується пилок. У пилках розвивається пилкове зерно - мікроспора, яка вкрита внутрішньою тонкою оболонкою (інтима) та зовнішньою (екзина). У процесі розвитку пилкового зерна шляхом мейозу, потім мітозу в ньому утворюється дуже редукований чоловічий гаметофіт, який складається з однієї вегетативної та однієї генеративної клітин. Генеративна клітина поділяється на два спермії, що беруть участь у заплідненні. Вегетативна клітина забезпечує життєдіяльність пилкового зерна та ріст пилкової трубки.
Насінні зачатки розвиваються на внутрішніх стінках зав'язі з горбочка, в закладанні якого беруть участь поверхневі шари клітин. Насінний зачаток зовні має один або два покриви - інтегументи, між якими залишається отвір - мікропіле.
Основу насінного зачатка називають халазою. Під інтегументом знаходиться багатоклітинний утвір - нуцелус, який гомологічний ме-гаспорангію. Одна з його клітин стає археспоріальною. З неї утворюються чотири мегаспори. Одна з мегаспор дає початок восьмиядерно-му зародковому мішку (жіночий гаметофіт). Після першого поділу два дочірні ядра прямують до протилежних полюсів гаметофіту і там діляться ще два рази. Троє ядер у кожного полюса утворюють по три клітини. Одна з клітин біля мікропіле стає яйцеклітиною, а дві інші - синергідами. Інші три ядра на протилежному полюсі утворюють клі-тини-антиподи. Решта двоє ядер мігрують до центра, зливаються, перетворюючись на вторинне диплоїдне ядро центральної клітини, яке дає початок ендосперму. Яйцеклітина в такому вигляді підготовлена до запліднення.
5. Суцвіття: біологічне значення, класифікація
У рослин пагін може закінчуватися однією квіткою або багатьма дрібнішими квітками, що утворюють суцвіття.
Суцвіття - це система видозмінених пагонів, які несуть квітки. Біологічне значення суцвіття полягає у кращому забезпеченні процесу запилення квіток (рис. 42).
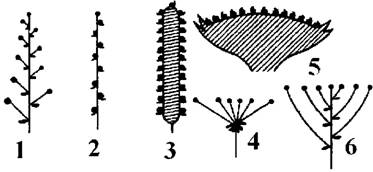
Рис. 42. Прості суцвіття (за Яковлевим, Челомбітько, 2001):
1 - китиця; 2 - колос;
3 - початок; 4 - простий зонтик;
5 - кошик; 6 - щиток.
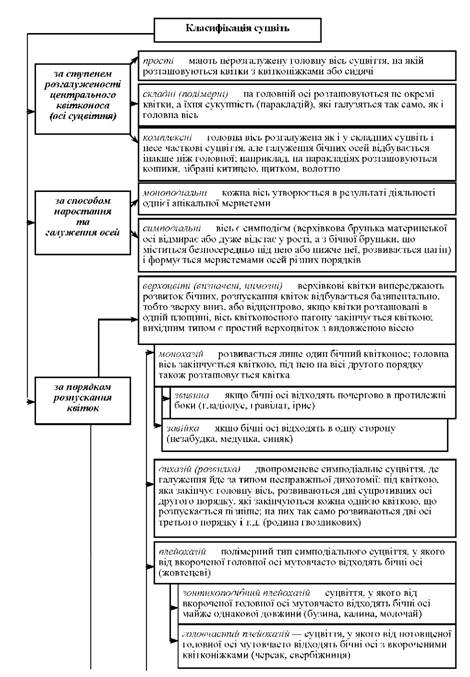
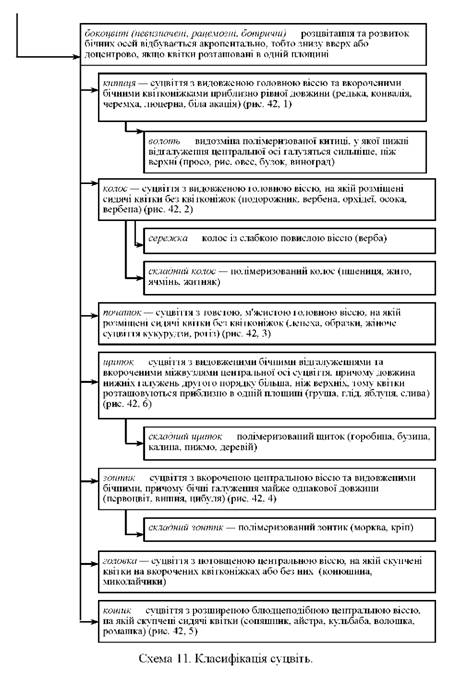
Класифікацію суцвіть подано на схемі 11.
6. Запилення та запліднення
Після того, як в пилкових мішках сформується пилкове зерно, стінки пиляків підсихають, розтріскуються, що зумовлює вивільнення пилку. Перенесення пилкових зерен на приймочку маточки називають запиленням. Типи запилення подано на схемі 12.
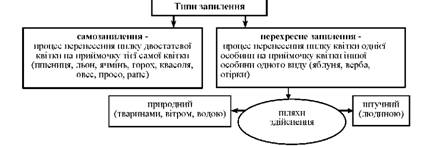
Схема 12. Типи запилення.
Спосіб запилення залежить від будови і фізіологічних особливостей квіток, зовнішніх умов тощо.
Запилення у самозапильних рослин відбувається ще тоді, коли квітки ще не розкрилися (арахіс, копитняк, розрив-трава, фіалка). Деякі самозапильні рослини здатні запилюватися і перехресно, але при цьому не утворюється потомство або потомство вироджується.
Особливості будови квітки самозапильних рослин:
1) у квітці є маточка й тичинки;
2) тичинки трохи довші за маточку.
Пристосування до перехресного запилення:
o самобезплідність (пилок квітки, потрапивши на маточку тієї ж квітки, не проростає);
o різночасове дозрівання тичинок і маточок у межах однієї квітки;
o здійснюється за допомогою вітру - анемофільні рослини (10%), або комах - ентомофільні рослини (90%), зрідка - води (гідрофільні), дрібних птахів (орнітофільні) (рис. 43), ссавців.

Рис. 43. Орнітофільна рослина - орхідея.
Особливості вітрозапильних та комахозапильних рослин подано в таблиці 16.
Таблиця 16
ОСОБЛИВОСТІ ВІТРОЗАПИЛЬНИХ ТА КОМАХОЗАПИЛЬНИХ РОСЛИН
Особливості Ознаки -1- вітрозапильних рослин комахозапильних рослин | ||
Місце зростання | на відкритих місцях, утворюючи зарості одного виду (очерет, ковила) | у різних кліматичних зонах континентів світу |
Час цвітіння напровесні, до розпускання лис- збігається з появою комах тя (ліщина, вільха, береза) | ||
Будова квітки | o квітки без нектарників, дрібні, малопомітні, із спрощеною небарвистою оцвітиною; o великі пиляки на довгих тичинкових нитках; o маточки відкриті з великими приймочками; o пилку утворюється багато; o пилок дрібний, легкий, з гладенькою поверхнею | o віночок яскраво забарвлений; o квітки поодинокі або зібрані у суцвіття; o на дні віночка утворюються нектарники з запашним солодким нектаром; o пиляки невеликі та утворюють порівняно з вітрозапильними менше пилку; o пилок великий, липкий, з шорсткою поверхнею |
Характерні ознаки гідрофільних рослин (кушир, валіснерія, стрілолист):
o пилок ниткоподібний, що забезпечує кращий контакт з приймочкою;
o утворення негігроскопічного пилку, який не гине у воді. Характерні ознаки орнітофільнихрослин (орхідеї) (рис. 43):
o квітки мають багато нектару;
o квітки мають яскраве забарвлення.
Біологічне значення запилення: від нього залежить подальше запліднення, розвиток плодів та насіння.
Біологічне значення перехресного запилення:
а) забезпечує обмін генами;
б) дає матеріал для природного добору (підтримує високу гетерозиготність популяцій);
в) зберігає стійке потомство - носіїв найбільш сприятливого співвідношення генів.
Штучне запилення застосовується людиною для:
а) підвищення врожайності (у холодну погоду, коли не літають комахи - запилювачі; у безвітряну погоду у вітрозапильних рослин; на виробничих посівах перехреснозапильних рослин);
б) отримання нових сортів рослин.
Після запилення відбувається процес запліднення - злиття чоловічої статевої клітини з жіночою:
1) пилок, що потрапив на приймочку маточки, продовжує свій розвиток;
2) вегетативна клітина пилкового зерна активізує ріст пилкової трубки;
3) за вегетативною клітиною в пилкову трубку переходить і генеративна клітина, з якої у результаті мітозу утворюються 2 спермії;
4) пилкова трубка за короткий час проходить крізь пухку тканину стовпчика маточки і проникає до зародкового мішка;
5) досягши яйцеклітини, пилкова трубка розривається, з неї виходять 2 спермії, а вегетативна клітина пилкової трубки руйнується;
6) один із сперміїв зливається з яйцеклітиною, утворюючи диплоїдну зиготу, з якої розвивається зародок нового рослинного організму;
7) другий спермій зливається з центральною /2п/ клітиною, у результаті чого утворюється триплоїдна клітина /3п/, яка дає початок ендосперму - запасу поживних речовин для зародка.
Вищеописаний процес був відкритий у 1898 р. професором Київського університету Святого Володимира (нині - імені Тараса Шевченка) Сергієм Гавриловичем Навашиним (1857-1930) і названий ним подвійним заплідненням.
Схематично цей процес можна зобразити так (схема 13):
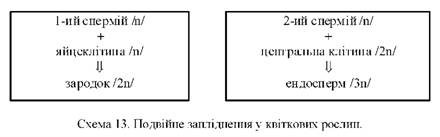
Біологічне значення подвійного запліднення полягає в тому, що один спермій зливається з яйцеклітиною, утворюючи диплоїдну зиготу, а внаслідок злиття другого спермія пилкової трубки з диплоїдним ядром центральної клітини зародкового мішка утворюється триплоїдний ендосперм, матеріал для живлення клітин зародку. Подвійне запліднення характерне тільки для покритонасінних і є новим кроком в еволюційному розвитку рослин.
Після запліднення із заплідненої диплоїдної яйцеклітини формується зародок насінини, а із триплоїдної клітини - поживна тканина (ендосперм), покриви насінного зачатка перетворюються в покриви насінини, а стінка зав'язі, розростаючись, утворює оплодень.
ЛЕКЦІЯ 11. Генеративні органи рослинного організму. насінина та плід
1. Насінина: загальна будова та функції
2. Особливості будови насінини однодольних та дводольних рослин
3. Плід та його біологічне значення
4. Класифікація плодів
5. Способи поширення плодів і насіння
Питання для самоперевірки
ЛЕКЦІЯ 12. Систематика рослин. нижчі рослини - водорості (algae)
1. Загальна характеристика нижчих рослин